
植物の栄養応答で機能するユビキチンリガーゼの基質認識機構を明らかに
形態機能学系・山口研究室の博士3年生・安田盛貴さんが論文を発表しました。以下、安田さんによる内容の解説です。
植物は光合成で得た糖と土壌中から吸収した無機窒素を,それぞれ炭素源(C)および窒素源(N)として,タンパク質の合成に利用します。したがって,生命活動を維持するためには,CとNの両者がバランスよく存在することが必要です。これを果たすために,植物は,細胞内のCとNのバランス(C/N)を感知し,それに応じて代謝・成長を制御しています。これはC/N応答と呼ばれ,生育を左右する重要な要素です。
私たちはモデル植物であるシロイヌナズナを用いて,C/N応答の分子機構解明に取り組んできました。これまでの解析から,ユビキチンリガーゼATL31が14-3-3タンパク質のユビキチン化とそれに伴うタンパク質分解を介して,C/N応答を制御していることを明らかにしました。しかし,ATL31がどのように14-3-3タンパク質のユビキチン化を制御しているのか,そのメカニズムは不明なままでした。
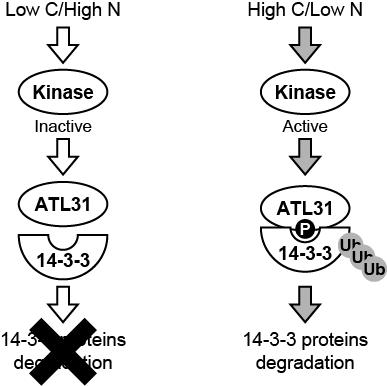
本研究では,ATL31のリン酸化が14-3-3タンパク質のユビキチン化制御に重要であることを明らかにしました。私たちはまず,ATL31と14-3-3タンパク質の相互作用解析およびATL31のリン酸化解析を実施しました。その結果,ATL31のC末領域に存在する4か所のセリン/スレオニン残基のリン酸化が,14-3-3タンパク質との結合に必須であることを突き止めました。そこで,これらのセリン/スレオニン残基を全てアラニンに置換した変異型ATL31の過剰発現株(35S-ATL31-4A)を作出し,その影響を解析しました。野生型(WT)に比べ,通常型ATL31過剰発現株(35S-ATL31)では高C/低N条件下における14-3-3タンパク質の蓄積が抑制され,生育阻害が緩和します。しかし,35S-ATL31-4Aはこのような表現型を示しませんでした(図1右側)。一方,低C/高N条件下では各植物体間で表現型に差はみられませんでした(図1左側)。これらの結果は,14-3-3タンパク質のユビキチン化を介したC/N応答制御には,ATL31のリン酸化が必須であることを示しています。また,35S-ATL31における14-3-3タンパク質蓄積の抑制は,高C/低N条件下でのみみられたことから,ATL31のリン酸化は高C/低N条件下では促進し,対照的に低C/高N条件下では抑制されていることが予想されます。今回の研究で得られた分子機構の概略を図2に示します。
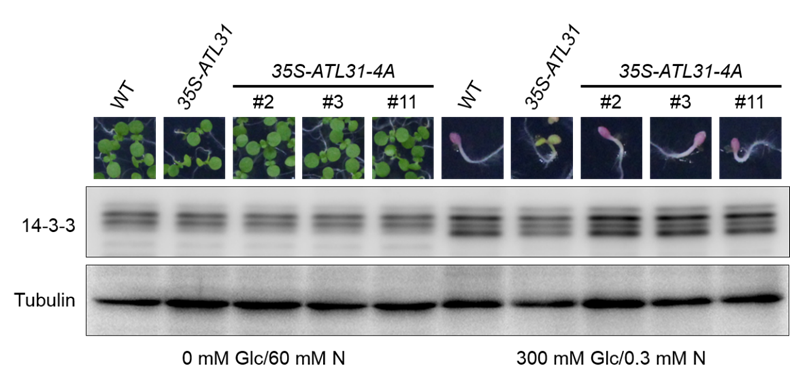
各植物体を低C/高N条件下(0 mM Glc/60 mM N)または高C/低N条件下(300 mM Glc/0.3 mM N)で生育し,発芽後成長と14-3-3タンパク質の蓄積量を解析した。35S-ATL31-4Aは独立した3つのラインを解析に用いた。Tubulinはローディングコントロールとして用いた。
Shigetaka Yasuda, Takeo Sato, Shugo Maekawa, Shoki Aoyama, Yoichiro Fukao, and Junji Yamaguchi (2014) Phosphorylation of Arabidopsis ubiquitin ligase ATL31 is critical for plant carbon/nitrogen nutrient balance response and controls the stability of 14-3-3 proteins. The Journal of Biological Chemistry 289: 15179-15193 http://www.jbc.org/content/289/22/15179.long